現在、我々の思考や記憶は、脳内の神経のネットワークを通る電流のパターンによって生じていると考えられています。
ある記憶を思い出すには、特定の神経ネットワークに電流を流せばいいということです。
でも、その前に、記憶を植え付ける=覚える作業があります。
この覚える作業は、神経ネットワークにおいて何を引き起こしているのでしょうか?
ヘブが予見した「記憶の痕跡」
ヘブ則とは 1949 年カナダの心理学者であった Donald Hebb が自らの著書『The Organization of Behavior』の中で唱えた仮説です。
これは記憶のメカニズムを考えるうえで、現在でも基礎的な概念として用いられる仮説です。
※ただし、この法則にそぐわない現象も認められているので、完全にすべてを説明する統一的な理論であるとはいえません。
同時に発火するニューロン間のシナプス強化
それをヘブは予言しました。
正確に言うと、ある2つのニューロン A と B が互いにつながっているとき、
A が B を発火させた、ということが繰り返し起こると
A から B へのシナプスの強化が起こる。ということです。それも、物理的にです。
今風に言えば、A から出たグルタミン酸を、B が受容体で受け止めるのですが、
その受容体の数が増えるといった現象で理解できます。
これは、記憶の痕跡がシナプスの物理的な変化として永続的に残るものであると考えられます。記憶は、目に見えない漠然としたものではなく、物理的なシナプスの変化として厳然と存在しているということなのですね。
もしこれが事実だとすれば、くり返し、同じことを学習すると、このようなシナプスの変化が起こることにより同じことを思い出しやすくなるということになります。
あるいは、何らかの動作の練習であれば、繰り返し練習するほど、その動作を行いやすくなっていくということです。
現在ではこの現象を長期増強(LTP)と呼んでいます。
ヘブ則に従う長期増強と、従わない長期増強のメカニズムがあるといわれています。 この記事では、ヘブ則に従う場合のメカニズムについてとりあげてみましょう。
なぜ、同時に発火したニューロンの間にそのようなことが起こるのでしょうか?
ヘブ則に従う長期増強のメカニズム
今ここに、シナプス結合している2つのニューロン A と B が存在するとして。
A⇒B というつながり。A が出力側、B が入力側。A から出た信号が B に入るものとします。
A も B も、それぞれ、いろいろなニューロンからの入力を受け取って、多数決で発火 するかどうかを決めています。つまり、閾値を超える信号量が入れば発火するので す。
もしこの A と B が同時に発火したとしましょう。
しかしそもそも、同時に発火したニューロン同士は、互いに同時に発火したということをどのようにして知るのでしょうか?
そのことを説明する前に、ニューロン内の電流は逆流するという事実について知っておく必要があります。
従来は、ニューロン内で発火が起こった時、それによって電流は一方通行に伝わると 考えられてきました。軸索という出力側へと流れていくということです。多分、これを読 んでいる皆さんもそう思っていると思います。
しかし、実は同時に逆流も起こしていて、入力側(樹状突起)へも電流が流れていくと いうことが発見されたのです。
すると、同時に発火した2つのニューロンの間で何が起こるのか、見てみましょう。 (1997 年に Markram らによって発見されたスパイクタイミング依存可塑性のメカニズ ム)
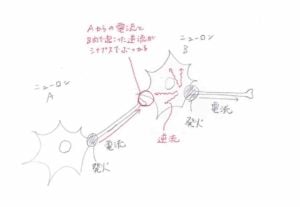
シナプス B が発火したとき、その電流は出力側(軸索方向)へと流れていくだけでな く、信号の入り口側にも逆流することがわかりました。したがって、A と B のシナプス 部にも逆流した電流が届きます。
すると、もし A と B が同時発火したならば、A からの信号を受けて B のシナプスから 起こる電流と、B 自身の発火から逆流した電流とが、A と B をつなぐシナプスに同時 に流れ込みます。
すると、普段の電流よりも大きな電流が流れることになります。
(具体的には、より多くのナトリウムイオンが流入し、より大きな電位で脱分極が起こり ます。)
この、「より大きな電位の脱分極」を感知するのが NMDA 受容体です。グルタミン酸受 容体の一種です。
この受容体は、「より大きな電位の脱分極」を感知すると、カルシウムイオンをニュー ロン B の中に流入させる、イオンチャンネルの役割を果たしている。
そして、カルシウムイオンが流入すると、ニューロン B の細胞膜の表面に、新たなグ ルタミン酸受容体が追加されるのです。このグルタミン酸受容体は、ニューロン A か らの信号をキャッチするためのものです。これが増えれば増えるほど、ニューロン A からの信号に対するニューロン B の反応は敏感になります。
ちなみに、A と B の発火のタイミングはほぼ同時か、わずかにずれたとしても、A が 先、B が後の順序になっていれば、シナプスは強化されます。
しかし、逆の順序に発火した場合は、かえってシナプス結合が弱まるのです。
だから、まったくランダムに発火しているだけでは、強化される確率と弱まる確率がほぼ半々で、変化が起こりません。
きちんと A⇒B という順序のパターンになったときだけ強化されるようになっているの です。
その他の長期増強のメカニズム
もっと古くから知られている長期増強のメカニズムとして 1973 年 Bliss と Lømo によ って観察された、高頻度の連続刺激による長期増強もあります。これも、やはり NMDA 受容体が反応することによって、シナプスにグルタミン酸受容体が増えるとい う結果に至ります。
ただ、NMDA 受容体を反応させるために用いる手段が、高頻度の連続刺激であると いう点が異なるだけです。
このメカニズムもまた、ヘブ則に従うものです。
しかし、ヘブ則に従わない神経可塑性のルールも発見されており、今後ますます研究 が進んでいくものと期待されます。
まとめ
長期増強は、学習の正体の一部であると考えられてます。 この記事で取り上げた長期増強のメカニズムは2つありましたが、いずれも NMDA 受 容体がそのカギを握っていました。NMDA 受容体が反応すれば、シナプスが増強され るのです。
そこで、NMDA 受容体を阻害したマウスで行った実験があります。学習できなくなって しまいました。
逆に、NMDA 受容体が通常より多いマウスをつくったら、記憶力が上がったのです。
もし、近い将来、記憶力や学習能力を増強するためのドーピング薬が開発されるとし たら、それは、この NMDA 受容体の数や感受性を高める作用をもつ薬なのかもしれませんね。

本日は、東京都世田谷ちゅうしん整体院 村山先生のコラムを紹介いたしました。
前回のコラム『自制心と頭の良さ:ワーキングメモリについて』こちら